|
| |
Evolution Encyclopedia Vol. 2 CHAPTER 15 – S P E C I E S E V O L U T I O N
"Throughout the past century there has always existed a significant minority of first-rate biologists who have never been able to bring themselves to accept the validity of Darwinian claims. In fact, the member of biologists who have expressed some degree of disillusionment is practically endless." —*Michael Denton, Evolution: A Theory in Crisis (1986), p. 327. "I personally hold the evolutionary position, but yet lament the fact that the majority of our Ph.D. graduates are frightfully ignorant of many of the serious problems of the evolution theory. These problems will not be solved unless we bring them to the attention of students. Most students assume evolution is proved, the missing link is found, and all we have left is a few rough edges to smooth out. Actually, quite the contrary is true; and many recent discoveries . . have forced us to re-evaluate our basic assumptions. " —*Director of a large graduate program in biology, quoted in Creation: The Cutting Edge (1982), p. 26. "Charles Darwin, himself the father of evolution in his later days, gradually became aware of the lack of real evidence for his evolutionary speculation and wrote: `As by this theory, innumerable transitional forms must have existed. Why do we not find them embedded in the crust of the earth? Why is not all nature in confusion instead of being, as we see them, well defined species?" — H. Enoch, Evolution or Creation, (1966), p. 139. Evolution is based on change from one species to another. In chapters 13 and 14, Natural Selection and Mutations, we have found that there is no mechanism by which it can occur, and In chapter 17, Fossils, we will learn that there is no past evidence of such change. But since the distinct plant and animal types are such a crux in the entire controversy, we will here devote a full chapter to speciation. This material will help fill out the picture of what we are learning in other chapters. ORIGIN OF LIFE CHAPTER 15 - SPECIES EVOLUTION BASIC ARRANGEMENT OF THIS CHAPTER Introduction Species have not and cannot change into different ones 1 - Classification systems 2 - Facts about species 3 - Disproving species evolution Appendices 1 - Origin of the species unknown 2 - The experts are puzzled 3 - Classifying plants and animals 4 - Micro and Macro 5 - Population genetics fails to prove evolution 5a - Lebzelter principle 5b - Hardy-Weinberg principle 6 - Cladists against evolution 7 - Millions of years for one species 8 - Our family tree 9 - The creature that fits no category Related studies: Chapter 13, Natural Selection Chapter 14, Mutations WHERE THE BATTLE IS—The battle over evolutionary theory finds its center in the species. This is where *Charles Darwin attempted to fight it, but without success: Why are there distinct species? *Darwin's theorized "evolving" should only have produced a confusion of life forms with no separate species. Why has cross-species change never occurred? Why can we not find how it could occur?
The problem of species has become a major unsolved problem of science.
In his book, Darwin never touched on the origin of the species, and the material he gave on the evolution of the species was totally inadequate.
For additional information see the appendix topic, "1 — Origin of the Species Unknown, " and " 2 — The experts are Puzzled" in the appendix. 1 - IDENTIFYING THE SPECIES PLANT AND ANIMAL CLASSIPICATIONS—The science of classifying plants and animals is called taxonomy. "Classification or taxonomy is the theory and practice of naming, describing, and classifying organisms. " –*W. Stansfield, The Science of Evolution (1977), p. 98. All plants and animals have been placed by taxonomists in logical categories, and then arranged on several major levels, which are these:
It should be kept in mind that there is no such thing as a phylum or a family. Those are just convenient names, and are like rooms in a zoo or botanical garden; each one with a different collection of plant or animal species. It is the species which are alive; the room is not. The terms "phyla, classes, orders, families," and most of the "genera" are merely category labels. It is only the species (with some genera included, which should be labeled as species) which count, for they only have any real, living existence.
Here is an example of how classification works. This is the classification of the house cat:
If you go to the zoo, you will see a sign on one cage, "Giant Panda," with the words, "Ailuropoda melanoleuca" just below it. The first line is the common name of this large black-and-white bear from China; the second line is its "scientific name." These two-part Latin names (called binomials or binominals) are understood by scientists worldwide. The first word is the Genus, and the second is species. Sometimes the name of the discoverer or namer is added as a third word. The Swedish naturalist, Linnaeus, invented this method of scientific nominclature in the 1750s. Darwin recognized that there was no evidence that any species had evolved from any other species. His contention that the only practical solution was, first, to classify plants and animals; second, point to similarities between them, and, then, declare that, therefore, one must have evolved from the other, or from a common ancestor. From beginning to end, evolution is just theory, theory, theory.
Little wonder few could understand it. They saw that Darwin was basically trying to prove an assumption (that species evolved from other species), with more assumptions.
THE GENESIS KIND—Back in the beginning, the law of the "Genesis kinds" was established:
In the same way, the birds, sea-life, and animals were each to reproduce "after their kind" (Genesis 1:20-22, 24-25). This principle was not to be violated. And this is what we find in the fossil record, and in the world today. The "Genesis kind" is generally equivalent to the species level, but sometimes the genus level. This variation is due to flaws in our humanly-devised classification systems. Since the Hebrew words used in Genesis for "create" and "kind" are bare and min, Frank Marsh, a careful research scholar in speciation, has suggested the term baramin as an identifying name for this "Genesis kind." (Min is used 10 times in Genesis 1, and 21 times in the rest of the Old Testament.) It would be a good word to use, since it is more accurate than "species," which can at times be incorrect. Another important term is the "biological species." BIOLOGICAL SPECIES—The term, "biological species," is increasingly becoming accepted as a basic reference point by scientists. Although there are instances in which obvious subspecies do not cross breed, biological species would normally apply to those species which do not cross-breed outside of their own kind. Here are some definitions of such a true species:
Marsh has studied the species question for decades. Here is his approach to trying to relocate the "Genesis kind:"
MICRO- VS. MACROEVOLUTION—Evolutionists point to changes within the species and call that "microevolution," and then proceed to tell us that such sub-species changes prove that there theorized changes across species (which they term "macroevolution") must also be occurring. But random gene shuffling within the species only produces new varieties and breeds. The DNA code barrier is not penetrated. Transformations across the species barrier never occur. New varieties end new breeds is not evolution; it is only variation within the already existing species.
COUNTING THE SPECIES— *Aristotle could list only about 500 kinds of animals, and his pupil *Theophrastus, the most eminent botanist of ancient Greece, listed only about 500 different plants. Through the centuries, as naturalists counted new varieties of creatures in the field, in the air, and in the sea, and as new areas of the world were explored, the number of identified species of animals and plants grew. By 1800 it had reached 70,000. Today there are several million. Two-thirds of them are animal and one-third are plant. The flowering plants and insects are the two largest single categories. JOHN RAY—John Ray (Wray) (1627?-1705) apparently was the first scientist to formerly recognize the "species." He prepared a large classification of all the species of plants and animals known in his time (about 18,600). Ray was an earnest Christian who, in the wonderful structures of plants and animals, saw abundant evidence of a Creator's hand. CARL LINNAEUS—Carl von Linne (1707-1778) spent his adult life as a teacher at the University of Uppsala. At the age of 50, he latinized his name to "Carolus Linnaeus. " The classification system of plants and animals developed by Linnaeus was to become the standard used today. He published it in his book, Systems Naturae in 1735. As was usual among scientists of his day, he used Latin descriptive names to identify each class and order, which were then subdivided into genera, and finally into species. As a result of his lifework of investigations into the entire field, Linnaeus came to two definite conclusions: (1) Species were, for the most part, the equivalent of the "Genesis kind." (2) There had been no change across the basic categories—now or earlier. As a result of his studies, Linnaeus arrived at a firm belief in Special Creation and the fixity of species. He said, "We reckon as many species as issued in pairs form the hands of the Creator" (quoted in *H.F. Osborne, From the Greeks to Darwin (1929), p. 187). Men today may call themselves experts in taxonomy, but it is significant that the two men in human history able to lay a solid foundation for biological classification—saw in all their findings only evidence of creation, not evolution.
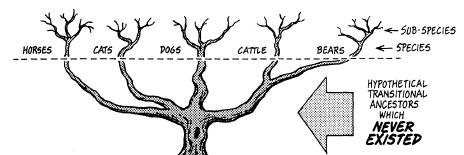
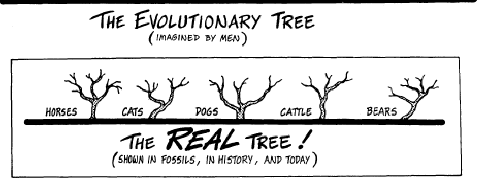
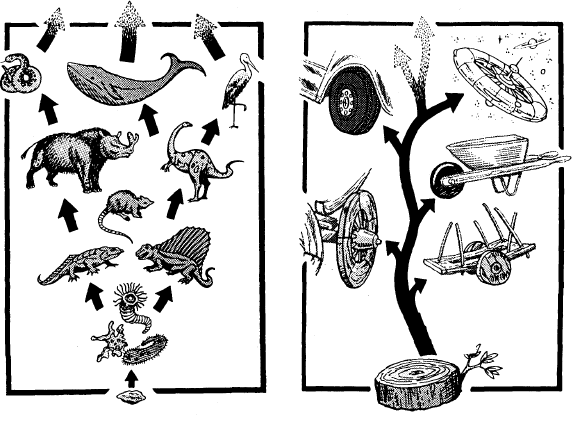
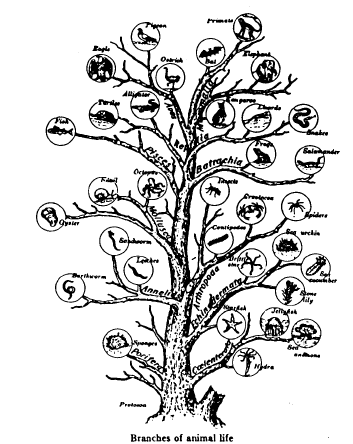
RAY AND LINNAEUS—Although the English naturalist Ray (1686) and the Swedish taxonomist Linnaeus (1738) were the first biologists to recognize the reproductive gap (the inability to cross) as the basic mechanism in nature which separates the true species, Linnaeus was the one to develop our modern system of classification. Unfortunately, he frequently listed as separate species, life forms which could interbreed. Some of these decisions were based on ignorance, but nevertheless we live with the results today. Thus, the true species are not always those which are listed in the textbooks as "species." It is now recognized by many qualified biologists that John Ray did better quality work, for he carefully adhered to biological species in preparing his species categories. In contrast, Linnaeus at times confused them by placing true species in genera or sub-species categories. THE TREE OF LIFE Reproduced below is a page from an old biology textbook. Notice the misleading wording: There is "constant progressive departure from ancestral types" and, "of course, only the main branches are shown." The textbook illustration only shows the twigs, because that is all there is! This diagram is intended to suggest the origin of various animal forms, with the constant progressive departure from ancestral typo, now in one direction and now in another, like the branching of a tree. Of course only the main branches are shown.
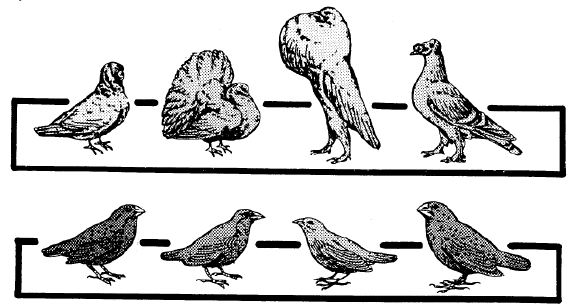
LUMPERS AND SPLITTERS—There has been a perennial problem in regard to the "Jumpers" and "splitters". There is a tendency for the taxonomists—the experts who classify plants and animals—to fall into one or the other of these two categories. The Jumpers place species together which should be divided into sub-species. The splitters tend to put true species into sub-species categories. "Lumper species," are also called "Linnaean species" because, back in the early 1700s, both Linnaeus and Ray pioneered the lumping of species. "Splitter species" are also called "Jordanian species" for the French botanist, Jordan, who initiated this approach in the early 1800s. So today we find both Linnaean and Jordanian species scattered throughout the scientific lists of plants and animals. It is important to keep this in mind, for selective breeding of Jordanian species can produce new species! This would appear to prove evolutionary claims, and indicate species cross-over as taken place—when, actually, two members of the same sub-species interbred. An example of such a splitter would be the taxonomist, *Walker, who lists seven species of cattle—all of which are in the same species. When the Santa Gertrudis cattle were developed in the 1960s by breeding zebu bulls with strains of Texas longhorns, Herefords, and shorthorns, the result was a new sub-species, but some splitters classify it as a "new species." Yet the Santa Gertrudis is merely another type of the cattle species, and able to cross-breed with several others. FAMILY TREE—Everyone has seen paintings in museums and textbooks of our "family tree," with its worms, birds, apes, and man shown in relation to how they evolved from one another. The impression is given that there can be no doubt that it really happened that way, for did not scientists prepare those charts? The truth is that the "Evolutionary Tree of Life" is just another fake, like all the other "evidences" of evolutionary theory. One example of what you will find on one "limb" of this imaginary "tree" are a mutually diverse group of creatures called the "coelenterates" solely because they have a saclike body, tentacles, and a single mouth opening. Although coral and jellyfish are not a bit alike, they are therefore classified together. We are supposed to believe that, because coral and jellyfish are together on the tree, one evolved from the other! In the plant kingdom, the Compositae is merely a waste-basket category that includes all the flowering plants that cannot be fitted in somewhere else. So therefore, they are supposed to have evolved from one another. This "tree" is a classificationist's nightmare! In chapter 21, Similarities, you will find a number of similarities never discussed by the classification experts. If used, they would produce totally different "tree" relationships. Two such examples would be aortic arch groupings and chromosome count comparisons. Evolutionists have tried to shoehorn species into various "families" of related species. But there are many creatures which have such unusual shapes, organs, or functions—that they fit into no particular pattern. The only reason the classification systems work as well as they do is because only a few surface distinguishing characteristics are used in the classification schemes. For additional information see the topics, a "8 - Our Family Tree, " and "9 - The Creature that Fits a no Category," in the appendix. 2 - FACTS ABOUT SPECIES INTERESTING FACTS ABOUT SPECIES—Here are some facts about species and subspecies that will help you understand some of the problems inherent in this Interesting field of plant and animal classification: 1 - Chickadees. The Carolina Chickadee (Parus carolinus) and the black-capped Chickadee (parus atrlcapillus) look just like each other in every way, and freely interbreed. Yet they have different songs! Although they have been classified as two different species, we have here one species with two alternate gene factors. 2 - Wheat. Linnaeus classified spring wheat (Trltlcum aestlvum L.) as a different species than winter wheat (T. hybernum L.). Yet they are both strains of the same wheat. They will cross and produce fertile hybrids. They should have been classified as sub-species. 3 - Ladybugs. The ladybird beetle (Coccinellidae) has been divided into a number of different "species," but solely on the basis of different wing covers and the number and arrangement of spots on their backs. 4 - Song sparrows. For over two centuries four species of sparrows in North America had been listed (lincoln, fox, swamp, and song). Gradually this number increased as taxonomists moved westward and found additional sparrows. Soon we had lots of sparrow "species." But as more and more were discovered, it was recognized that they were but intermediates between the others! So the experts finally got together and reclassified them all as sub-species of but one species, the song sparrow (Passerella melodia). 5 - Horses. The horse (Equus caballus L) and the zebra (E. zebra L.) were both classified by Linnaeus as different species, yet they are only varieties of the horse kind, and are generally cross-fertile. 6 - Foxes. The red fox (Vulpes fulva) and the Newfoundland red fox have been categorized in different species, although the only difference is a paler reddish coat and shorter tail for the Newfoundland variety. Six taxonomists list 10 varieties of red fox, while 2 others list one species (Vulpes fulva) and count 12 sub-species. 7 - Cattle. There are several different subspecies of cattle (Bos taurus L.). Although the American bison (Bison bison L.) and the European bison (Bison bonasus L) have a similar morphology (appearance), they will still generally crossbreed with cattle. In addition, it has been discovered that the African buffalo (Syncerus Gaffer) also interbreeds with them—yet the bison and cattle have been placed in totally different genera. 8 - Corn. One expert (*Sturtevant) categorized 6 species of corn (sweet, flint, flour, pod, dent, and popcorn), while other taxonomists acknowledge that they are all only varieties of one species. 9 - Finches. In the chapter on Natural Selection and History of Evolutionary Theory, we discuss *Charles Darwin's finches (13,14,17, or 19; the count varies regarding this look-alike bird), which he found on the Galapagos Islands. Although about the same in size, shape and color, and together form a set of subspecies of finches which originally came from South America, yet Darwin called them different species—and therefore a proof of evolution. Those finches made a strong impression on his mind. INCREASING SUBSPECIES—Many different sub-species exist for some life forms, while there are but few for others. A key factor seems to be the ability of the creature to travel, whether by seed, spore, or in person. For example, the tiny fruit flies cannot travel very far, so there are many varieties of them. The animal with the most sub-species appears to be the southern pocket gopher (Thomomys umbrinus) with 214 subspecies, and, next to it, the northern pocket gopher (T, talpoides) with 66. Another highly-isolated species is the deer mouse (Peromyscus maniculatus), with 66 subspecies. In the case of animals which have been domesticated, such as dogs, cats, cattle, sheep, pigeons, and chickens, there are many sub-species as a result of selective breeding. The same holds true for cultivated crops (corn, beans, lettuce, cabbage). It was indeed strange that sub-species got *Darwin off on the wrong track.
There are instances in which sub-species generally do not breed across sub-species. The other extreme is instances in which animals above the species level will breed and apparently have young. In some cases these are true species, and should have been classified as such. But there are also instances in which breeding did NOT occur—although it appeared to take place! In true fertilization, the male and female elements unite and produce young. But there are times when non-species have been bred and young have been produced-in which no true breeding occurred! This false breeding takes place when the presence of male sperm stimulates the egg to begin production on a new life form, but the sperm is rejected because it is from a different species. The resulting birth is known as parthenogenesis. Scientific analysis has established that this false breeding across true species works in exactly the manner described here. It is significant that mankind can never successfully breed across with any other species, including any of the great apes.
That which evolutionists call "microevolution," is nothing more than breeding within species and producing varieties (subspecies); it is not evolution. Men have bred certain domesticated animals for thousands of years, but all they have produced is variations of those animals—nothing else. For example, a cow may produce an Brown Swiss, a Devon, an Africander, a Holstein-Friesian, an Aberdeen-Angus, a Jersey, a Brahman (Zebu), a Galloway, a Shorthorn, a Guernsey, a Hereford, or a Catalo. But they are all merely varieties of cattle. Producing new breeds in animals is not evolution; producing new varieties in plants is not evolution. One careful researcher (Frank Marsh) spent years tracking down every report of crosses above that of true species. Each time he found them to be hoaxes. One instance was of bird feathers sewn to a stuffed animal skin. It made good copy for a newspaper article, so it was printed. 3 - DISPROVING SPECIES EVOLUTION MENDELIAN GENETICS—It has been said that the foundations of evolutionary theory were laid by the work of *Charles Darwin (1809-1882), but that the principles which Gregor Mendel (1822-1884) discovered, as he worked with garden peas at about the same time that Darwin was writing his book, were the means of abolishing that theory. Everyone is acquainted with the illustration of the rough and smooth-coated guinea pigs. It was the work of Mendel that formed the basis for understanding the transmission of inherited characteristics. Mendel prepared the foundation for modern genetics. It was later discovered that within the cell are chromosomes, and inside the chromosomes are genes, and inside them is the coded DNA. (For more information on this, see chapter 10, DNA.) Random shuffling of the genetic code is what determines whether or not that baby guinea pig will inherit a rough or a smooth coat from its parents. But either way he will remain a guinea pig. Because that tiny newborn creature is locked into being a guinea pig is the reason why Darwin's theory crumbles before the science of genetics. PRIMITIVE ANCESTERS—Evolutionists tell us that certain creatures are more "primitive" than others, and are their "ancestors." But that is just theory. Consider but one example: the monotremes and the marsupials, which are supposed to be "primitive ancestors" of the mammals:
MANY VARIATIONS POSSIBLE—Yes, variations are limited by the species barrier—but how many potential variations are possible within a given species? This is an important question. Francisco Ayala has calculated that, among humans, a single couple could theoretically produce 102017 children before they would have to produce one that was identical to one of their earlier children (not counting identical twins, which came from the same egg and sperm). That would be 1 followed by 2,017 zeroes. The number of atoms in the known universe is only 108. So the number of possible variations within any given species is quite broad. Yet all of them would only be variations within the same species. ALWAYS A LIMIT—Then there is artificial selection. We discussed this at length in chapter 13, Natural Selection, and found it to be highly selective plant and animal breeding. In regard to any given single factor, selective breeding may be carried out, but soon a limit in factor variety will be reached. What limits it? It is the DNA code in the genes. That code forbids a crossover to a new species. The genetic makeup within the chromosomes forms a barrier; a literal wall of separation between one species and another. (For several scientific quotations regarding this, see the selective breeding quotation section at the back of chapter 13, Natural Selection.)
LIMITS OF VARIABILITY—This IS a crucial factor. All evolutionary theory pivots on whether or not there are such limits. Can one species change into another one? If there are definite limits forbidding it, then evolution cannot occur. An evolutionary encyclopedia provides us with a brief overview of the history of theory and "pure—line research" into limits of variability:
Whatever we may try to do within a given species, we soon reach limits which we cannot break through. A wall exists on every side of each species. That wall is the DNA coding, which permits wide variety within it (within the gene pool, or the genotype of a species)—but no exit through that wall.
LOSS OF FITNESS—So then, a limiting wall will always be reached, but the variations made within those borders do not actually bring overall improvements in the corn, cows, and chickens. All of the apparent improvement is made at the expense of overall fitness for life. Gish explains why this is so:
GENE DEPLETION—The scientific name for this loss of fitness through adaptation is gene depletion. According to this principle, selective breeding always weakens a species—and never strengthens it.
In chapter 14, Mutations, we deal with the generic load, that is mentioned in the above quotation. The original stock was strong, but as it branched out into variations within its kind, it became weakened. That is gene depletion. In addition, with the passing of time, genes are damaged through random radiation and mutations occur. Such mutations are always weakening, and gradually a genetic load is built up. Thus we see that, on one hand, the farther the species strays from its central original pattern, the weaker it becomes (gene depletion). On the other, as the centuries continue on, mutational weaknesses increase in all varieties of a given species (genetic load). The total picture is not one of evolving upward, strengthening, improving, or changing into new and diverse species. EVOLUTION WOULD WEAKEN AND NARROW—It is an astounding fact that evolutionary theory, if true, could only produce ever weaker creatures with continually narrowed adaptive traits. Consider this:
Well, that is a breath-taking discovery! If we had actually descended from monkeys, then we would have less genetic potential than they have! Our anatomy, physiology, brains, hormones, etc. would be less competent than that of a great ape. In turn, the monkey is supposedly descended from something else, and would therefore have less genetic capacity than its supposed ancestor had. Somewhere back there, the first descendent came from protozoa. All that follows In the evolutionary ladder would have to have considerably less genetic potential than protozoa! How can evolutionary theory survive such facts! Evolution ranks as one of the most foolish ideas of our time, yet it has a lock-grip on all scientific thought and research. The theory twists and warps all conclusions in an effort to vindicate itself. Just imagine how much farther along the path of research and discovery we would have been if, a hundred years ago, we had throttled evolutionary theory to death. SELECTIVE BREEDING—Selective breeding occurs when people thoughtfully select out the best rose, ear of corn, or milk cow, and then through careful breeding, produce better roses, corn ears, or milk cows. But please notice several facts in connection with this: (1) "Selection" requires intelligence, planning, and consistent effort by someone who is not the rose, corn, or cow. Random action is not "selection." Therefore "natural selection" is a misnomer. It should be called "random activity." (2) Contrary to what the evolutionists claim, selective breeding can provide no evidence of evolution, since it is intelligent, careful, planned activity, whereas evolution, by definition, is random activity. (3) Although random accidents could never produce new species,—neither can intelligent selective breeding! Selective breeding never, never produces new species. But if it cannot effect trans-species changes, we can have no hope that evolutionary chance operations could do it. (4) Selective breeding narrows the genetic pool, and although it may produce a nicer-appearing rose, at the same time it weakened the rose plant that grew that rose. Selective breeding may improve a selected trait, but tends to weaken the whole organism. Because of this weakening factor, national and international organizations are now collecting and storing "seed banks" of primitive seed. It is feared that diseases may eventually wipe out our specialized crops, and we need to be able to go back and replenish from the originals: rice, corn, tomatoes, etc. POPULATION GENETICS—A related area is termed population genetics, and it is declared by evolutionists to be another grand proof of their theory. Population genetics looks at locations of species and variations within a species, and theorizes evolutionary causes and effects. This field of study includes analysis of: (1) "Geographic isolation" of species, and sub-species produced by that species while in isolation. Some of these sub-species may eventually no longer interbreed with related sub-species, but they are obviously closely-related sub-species. (2) "Migration of populations" into new areas resulting occasionally in permanent colonization. Additional sub-species are produced in this way. (3) "Genetic drift" is analyzed. This is the genetic contribution of a particular population to its offspring. Variability here arises primarily from normal gene reshuffling. It is because of gene reshuffling that your children do not look identical to you. This is quite normal, and does not make your children new species! Population genetics, then, is the study of changes in sub-species. The information produced is interesting, but it provides no evidence of evolution, because it only concerns sub-species. For additional information see the appendix topic, "5 - Population Genetics." A field closely related to population genetics is selective breeding of plants and animals. But a favorite study of the population geneticists is people. Human beings are all one species. Population genetics analyzes changes within the "people species." Yet changes within a species is not evolution; only changes across a species constitutes genuine evolution.
A chapter closely related to population genetics is the one dealing with Geographic Distribution (chapter 27). You will also want to study the material on species chromosome counts, to be found in chapter 21 (Similarities). It will provide you with another clear disproof of evolutionary theory.
The common pigeon occurs in a remarkable number of varieties. Yet they are all pigeons, and every biologist acknowledges them as such. They are all members of the same species. Yet Darwin's finches, which vary but little, are said by some taxonomists to represent 14 different species!
GENETIC DRIFT—"Genetic Drift" is frequently spoken of as another "evidence" of evolution, but even confirmed evolutionists admit it proves nothing in regard to evolution. Genetic drift is changes in small groups of sub—species that have become separated from the rest of their species. Oddities in their DNA code factors became more prominent, yet there was no change in species.
You can see from the above statement that all that "genetic drift" refers to is changes in a "subspecies" of a plant or animal (or in a "race," which is a sub-species among human beings). Even *Rhodes recognizes that genetic drift provides no evidence of change from one species to another. This is due to the fact that all the drift has been found to be within species and never across them. The DNA species barrier forbids such traps-species changes. THE MALE/FEMALE REQUIREMENT—Inherent in the species quandary is the male and female element problem. It would be so much easier to bear young and, hopefully, produce new species, if everyone were females. But because it requires both a male and female to produce offspring, any possibility of going traps-species would mean producing not one new creature but two! Only recently was the extent of this problem fully realized.
There are well over three dozen different, distinct subspecies of dogs in the world. Yet they are universally acknowledged by scientists to be but members of the one dog species.
ALTERNATE ORIGINS OF THE SPECIES—Because of the inflexible nature of the species, *Austin H. Clark, a distinguished biologist on the staff of the Smithsonian Institution, wrote a shocking book in 1930. He had concluded that, since there was no evidence now or earlier of any crossovers between species—all of the major groups of plants and animals must have independently originated out of dirt and sea water!
But *Clark went further in his analysis. Ignoring the fossil evidence indicting no transitional forms but only gaps between species, he said that separate evolutions and origins had to have occurred—for there were simply too many differences between the various life forms. They could not possibly have evolved from each other. *Clark's book shook up the scientific world. The evolutionists tried to quiet matters, but about a decade later, ' Richard Goldschmidt published a different alternative view: gigantic million-fold mutations must have occurred all at once, that suddenly change one species to another. Goldschmidt's dreamy theory is today becoming more accepted by evolutionists, under the leadership of *Stephen Jay Gould. *Clark recognized the impossibility of evolution across major groups of plants and animals. Therefore he said each one independently originated out of sand and seawater. *Goldschmidt and *Gould recognized the impossibility of evolution across species, so they theorized that once every 50,000 years or so, a billion positive, cooperative, networking mutations suddenly appeared by chance and produced a new species. (For more on this, see chapter 14, Mutations.) THE CLADISTS—What about the experts in classifying plants and animals; what do they think about all this controversy over species and ancestral relationships? Scientists who specialize in categorizing life forms are called taxonomists. A surprising number of them have joined the ranks of the cladists. Cladist comes from a Greek noun for "branch." Cladists are scientists who study biological classifications solely for its own sake—for the purpose of discovering relationship, apart from any concern to determine ancestry or origins. In other words, the cladists are scientists who have seen so much evidence in plants and animals that evolution is not true, that, as far as they are concerned, they have tossed it out the window and instead simply study plants and animals. They want to know about life forms because they are interested in life forms, not because they are trying to prove evolution. Cladists are biological classification specialists who have given up on evolution. They recognize it to be a foolish, unworkable theory, and they want to study plants and animals without being required to "fit" their discoveries into the evolutionary "anteater" and "descendent" mold. They are true scientists; concerned with reality, not imaginings.
For additional information see the appendix topic, "8 - Cladists Against Evolution." THE SPECIES ARE NOT CHANCING—If one species cannot change into another, there can be no evolution. But this should not be surprising, for the fossil record reveals that the bat has not changed since it first appeared in the fossil record, supposedly "50 million years ago,"—and there was no transitional form preceding it. The same can be said for the other creatures. Throughout the fossil record, there are only solid, fixed forms, and wide gaps between. Those gaps are no surprise to us, but they are agonizing for the evolutionists. In chapter 17, Fossils, we go into detail on such matters
All this is a most terrible problem for the evolutionists.
Evolutionists have reason to be troubled: all the evidence they can find to substantiate their claims is changes within species (so-called "micro-evolution," which is not evolution), never changes across species ("macro-evolution," which is evolution).
The speciation problem is a gap problem. There are no transitional species, as there ought to be if evolution was true. In fact, there should not be any distinct species at all. (If evolution was correct; there would only be innumerable transitions) But, in great contrast, we find that there are absolutely no transitional forms to fill the gaps. In desperation, evolutionists have come up with an answer: "the transitions were made so slowly that they left no remains behind." —Wait a minute! How can that be? The more slowly the transitions, the larger would be the number of transitional forms which would be in the fossil strata for posterity to examine!
—And none other than *Charles Darwin himself agrees with us! "When we descend to details, we can prove that no species has changed [we cannot prove that a single species has changed]; nor can we prove that the supposed changes are beneficial, which is the groundwork of the theory." —*Charles Darwin, in "Francis Darwin (ed.), The Life and Letters of Charles Darwin Vol. 2 (1887), P. 210. How can there be millions of species, when the evolutionists tell us it takes a million years just to make one species? At that rate, in a billion years only a thousand could have been made! For additional information see the appendix topic, "7- Millions of Years for One Species." SUB-SPECIES CONTINUE TO BE PRODUCED—Evolutionists require long ages for each species to be produced. For example, Zeuner explains that the quickest pace that evolution can turn out new species is only one in every a 500,000 years.
But evidence for new species does not exist. Although new species have not been made before and are not being made now, sub-species are continually being produced. This is done primarily by gene reshuffling. And this making of new sub-species occurs with surprising rapidity. When isolated for several years, they sometimes no longer breed across sub-species—yet they are still sub-species and not different species. Here are some examples:
The above facts explain why there are such an abundance of so-called "species" in the world today. In reality, large percentage of them are just sub-species.
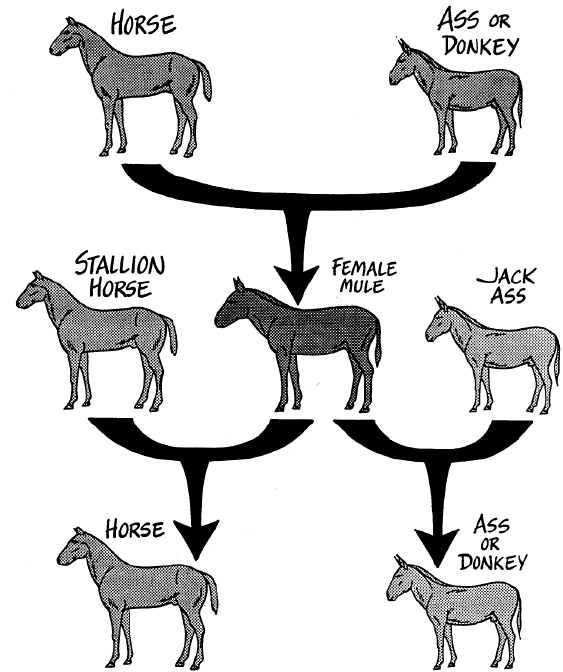
The horse is a distinct species and the donkey is a distinct species. A stallion horse bred with a female donkey will produce a hinney, which is small and not too strong. A male donkey mated with a mare horse will produce a mule, which is larger, stronger, and has more endurance than the donkey, but retains its sure-footedness and braying voice. From its mother, it inherits a large, well-shaped body and strong muscles, as well as a horse's ease in getting used to harness. From the donkey it also receives the quality of saving its strength when it has to work hard and for a long time. Mules also resist disease well. Properly cared for, mules can do as much work as horses, but under harder conditions. Mules are half-way between two species, so are sterile. Rarely does a female mule give birth, but when that happens it is because the mule was bred to a male horse or donkey. In such cases, the offspring will be three-fourths horse or donkey; it will not be a mule.
There is far too much jumbling of sub-species with species by the taxonomists. The word "species" is frequently used by scientists in a loose sense to include a multitude of sub-species.
In contrast, we have the fact that the subspecies are simply that and no more. All the Australian rabbits brought from Europe are only variations of the European rabbit. Both naturalists and paleontologists agree that the basic life forms—the true species—do not change.
Pages of quotations from evolutionary scientists will be found in the back of chapter 17, Fossils and Strata, attesting to the truth that there are only gaps between the species in the fossil record. THERE SHOULD BE NO SPECIES—As mentioned earlier, there ought to be no species at all Categories of plants and animals can be arranged in orderly systems only because of the separateness of the species. But if evolutionary theory was correct, there could be no distinct species. Instead, there would only be a confused blur of transitional forms, each one different than the others.
A leading evolutionist cannot understand why there are any species at all.
On one hand there is immense complexity within each species, but an a distinct barrier between species.
*Taylor then goes on to cite two great species problems: (1) If evolutionary theory be true, then why do separate species exist at all? (2) Why are living species identical to those alive "millions of years ago"? Here is his comment on the first of these:
But *Taylor's second problem is merely an extension of the first: (1) Why are there distinct species today? (2) Why do these distinct species extend all the way back as far as we can trace into the past?
SPECIES NOT CHANGING—So we have this third problem: If evolution is constantly producing species, why are the species not changing?
SUB-SPECIES RUNNING WILD—There is abundant evidence that additional sub-species are rapidly being produced. Hundreds of subspecies of cichlids have been identified in certain African lakes. The gene pool of the one cichlid species was large enough to allow for these many sub-species variations. (*Taylor, below, refers to them as "species," but they are really subspecies.)
THE REPRODUCTION PROBLEM —Evolutionary theory does not need sexual reproduction, yet we have it. How could two of each species-independent of each other—evolve? Yet this is what had to happen. The male and female of each species are forever uniquely separate from one another in a variety of ways, yet perfectly matching partners would have had to evolve together, at each step. Evolution cannot explain this.
DARWIN'S BEQUEST—It is well-known that *Charles Darwin had little to say about the actual origin of the species—the origin of life in a "primitive environment," but, instead, focused his entire work on an attempt to disprove fixed species. Yet, with the passing of the years, he became so confused regarding the species question that he was no longer certain how species could possibly change into one another. In his will, he gave a bequest to the Royal Botanic Gardens at Kew, England, which was trying to prepare the Index Kewensis, a gigantic plant catalogue which would classify and fix all known plant species.
Ironically, *Charles Darwin's last act was money given to help categorize the separate species. CONCLUSION—Here is how one author ably summarized the situation: "Anyone who can contemplate the eye of a housefly, the mechanics of human finger movement, the camouflage of a moth, or the building of every kind of matter from variations in arrangement of proton and electron,—and then maintain that all this design happened without a designer, happened by sheer, blind accident—such a per son believes in a miracle far more astounding than any in the Bible. "To regard man, with his arts and aspirations, his awareness of himself and of his universe, his emotions and his morals, his very ability to conceive an idea so grand as that of God, to regard this creature as merely a form of life somewhat higher on the evolutionary ladder than the others,—is to create questions more profound than are answered." —David Raphael Klein, "Is There a Substitute for God?" in Reader's Digest, March 1970, p. 55. For additional information see the appendix topics, "5A — The Lebzelter Principle, " and "5B – The Hardy-Weinberg Principle." POSTSCRIPT: SOON THEY WILL BE GONE—Interestingly enough, although the evolutionary problem is that the species are not changing, mankind's problem is that the species are disappearing!
"Within the past 40 years, rain forests in Brazil, Indonesia, Central America and Hawaii have shrunk from 5 million to 3.5 million square miles. More than 25,000 square miles are lost each year. If these forests disappear, as they will if destruction goes unchecked, thousands of species of plants and animals will disappear forever before they have even been discovered, named or studied." —*R. Milner, Encyclopedia of Evolution (1990), p. 383. "As the human population shoots up to 10 billion during the next century, the fate of thousands of species is sealed. Since man has appeared on Earth, we have increased the rate of extinction of other species by 1,000 percent." —*Op. cit., p. 202.
You have just completed S
P E C I E S E V O L U T I O N
|